
Increased body fat is a species specific characteristic of human infants. Compared to most other, non-aquatic animals, we give birth to infants with relatively amounts of body fat. This has been hypothesized to explain why our infants can sustain a weight loss of 8-10% after birth while receiving immune factor rich colostrum. Infant body fat frequently increases across the first few months of life, and it is generally thought that this is in anticipation of nutritional stresses that may be associated with weaning and independent motor development (crawling, walking).
However, baby fat is not all the same – babies actually have multiple different types of fat (also called adipose tissue). The most common form of adipose tissue we think about is white adipose tissue (think about the line of fat on a piece of pork or steak) and infants certainly have plenty of this white adipose tissue (WAT). But infants, since they can’t regulate their body temperature like adults can nor can they shiver, also have brown adipose tissue (BAT) and beige adipose tissue (BeAT). Brown and beige adipose tissue are involved in helping the infant to maintain body temperature, especially when exposed to colder temperatures. BAT and BeAT are able to do this because the individual cells are filled with mitochondria – organelles within the cells that produce heat (flash back to high school biology – mitochondria are the “powerhouse” of the cell and produce energy through ATP-synthesis). WAT cells, by comparison, contain very few mitochondria. When an infant is exposed to cold temperatures, the BAT and BeAT cells activate their mitochondria and start producing heat. This heat production helps keep babies warm in cold temperatures.

As infants age, BAT and BeAT typically decline (at least in the studies to date) and it was long thought that by adulthood, BAT was virtually absent (Carey and Kingwell 2013). We now know that BAT persists through childhood (Gilsanz et al., 2013), and into adulthood (Rogers et al., 2015), and may also vary by climate exposure – that is if you live in a cold climate like Siberia, you may maintain more BAT into adulthood than if you live in Columbia (Levy et al., 2018). Again though, the difficulty of studying BAT and BeAT has made investigations into global population variation in BAT quite difficult to study but new techniques using thermal imaging cameras promise new research (Levy, 2019).

When it comes to BAT, BeAT, and babies, we actually know very little about the production and maintenance of BAT and BeAT (this is actually true for all ages). Human infants are born with more WAT and BAT than most other primates and many mammals, but how these tissues are maintained has been largely unknown. In adults, there is some argument that cold stress and exercise may contribute to BAT retention into adulthood (ref), but babies clearly aren’t running marathons and what can explain the differences seen in rates of BAT and BeAT decline in different infants based on the handful of studies?
The answer may be BREAST MILK. I called this in 2013. Well, actually a community member in my longitudinal milk study in Nepal called this. He mentioned, one freezing night at 13,000 feet as we sat shivering by the fire that the babies in his community were different – they could handle the cold better. He asked me if it might be something in the breast milk. I’ve spent the last 6+ years trying to answer this question and am delighted to say the answer appears to be YES (although the specifics for this and other high altitude communities may differ).
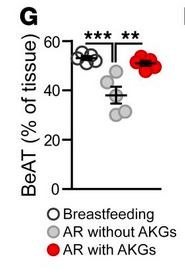
Which brings us to this amazing new paper by Yu and colleagues (full citation at the end; you can also view a video from the researchers HERE) looking at the role of breast milk and BeAT maintenance. This is an animal model study – the authors used mice to look at BeAT maintenance in a living model. However, this is coupled with subcutaneous fat biopsy data from 20 infants ages 2 to 12 months, half breastfed and half formula fed. Infants receiving breast milk had more BeAT in their subcutaneous fat than infants who had never received human milk or had only been breastfed for a short time. A key hormone, UCP1, associated with BeAT was largely absent in the tissue sample from infants not receiving breast milk but peaked at 2-4 months in breastfed infants. They were even able to show a dose effect – infants receiving breast milk for 30% or less of their lives had low or absent levels of UCP1 in these tissues AND lower levels of BeAT. When the infants were matched by age, formula fed infants had much lower BeAT and UCP1 than breastfed infants – if UCP1 was detectable at all.

In the mice, the researchers looked at targeted supplementation of newborn mice. The researchers had hypothesized that a special group of nutrients in human milk, alkylglycerols (AKGs), might be responsible for the maintenance of BeAT in human infants. Human milk is rich in AKGs, as is mouse milk while cow milk contains almost no AKGs. The AKGS in human milk are chimyl alcohol, batyl alcohol, and selachyl alcohol (none derived from alcohol that people drink; this is a chemical term used to describe their structure).

In infant mice, blood levels of AKGs were very similar to milk levels of AKGs, suggesting that milk was the source of AKGs for the infant mice. Mice pup blood levels of AKGs could be increased by feeding mice moms AKGs, which increased AKGs in her milk and her pups’ blood. To test the association between AKGs in milk and infant BeAT, the researchers fed one group of mice moms supplemental AKGs in their diet and another group of mice moms the same diet without any AKGs. The diet was fed for the first half of the mice pups’ infancy (days 3-10; mice nurse for about 21 days), a time period established to be a critical period in the development of WAT in mice.
The pups receiving AKG enriched mouse milk had reduced WAT and increased BeAT, but no change in lean tissue mass, at weaning compared to the pups receiving the AKG deficient mouse milk. This study was then followed by another mouse model, this time using artificial milk. Three groups of mice were used: controls who were mice pups who stayed with their birth mothers and received her milk, and two groups of mice pups fostered by non-lactating mouse mothers who received artificial milk – one containing AKGs and one group receiving milk not containing AKGs. The group of mouse pups receiving AKG supplemented formula were more similar in BeAT and WAT levels at weaning to the mouse milk control group than to the non-AKG group, suggesting that AKGs contributed significantly to the differences in BeAT observed between the groups (but not exactly the same).
Breast milk AKGs (and likely other hormones as well) directly contribute to the maintenance of BeAT and BAT tissues in human infants! This is really cool, as AKGs are a normal part of human breast milk (we don’t know much about population variation) produced during lipid secretion in the breast. Cows also have this mechanism of producing lipids for milk, but their milk contains specific micro-RNA that actually inhibit BeAT development – so formula is functionally absent of both the BeAT promoting AKGs and contains the BeAT inhibiting micro-RNAs.
Where do we go from here? So much more work is needed to understand AKG variation in human milk and how that may reflect differences in the ecological niches for infants. I would predict that milk from mothers in cold climates might have higher levels of AKGs to promote more BeAT and BAT in infants, while infants living in tropical climates may need less AKGs. Interestingly, even looking at patterns of infant feeding and milk composition across populations, without knowledge of their AKG content, provides some preliminary evidence in support of this – frequent nursing and more dilute milks commonly reported in hot climates should contain less AKGs than the milk of mothers nursing less frequently and producing higher fat milks in colder climates. It is just another amazing example of how human milk is shaping infant biology in response to environmental cues.
References
Carey AL, Kingwell BA. (2013) Brown adipose tissue in humans: therapeutic potential to combat obesity. Pharmacol Ther. 2013 Oct;140(1):26-33. doi: 10.1016/j.pharmthera.2013.05.009
Gilsanz V, Hu HH, Kajimura S. (2013) Relevance of brown adipose tissue in infancy and adolescence. Pediatr Res. 2013 Jan;73(1):3-9. doi: 10.1038/pr.2012.141.
Levy SB, Klimova TM, Zakharova RN, Federov AI, Fedorova VI, Baltakhinova ME, Leonard WR. (2018) Brown adipose tissue, energy expenditure, and biomarkers of cardio-metabolic health among the Yakut (Sakha) of northeastern Siberia. Am J Hum Biol. 2018 Nov;30(6):e23175. doi: 10.1002/ajhb.23175.
Levy SB. (2019) Field and laboratory methods for quantifying brown adipose tissue thermogenesis. Am J Hum Biol. 2019 Jun 4:e23261. doi: 10.1002/ajhb.23261.
Rogers NH. (2015). Brown adipose tissue during puberty and with aging. Ann Med. 2015 Mar;47(2):142-9. doi: 10.3109/07853890.2014.914807.
Yu H, Dilbaz S, Coßmann J, Hoang AC, Diedrich V, Herwig A, Harauma A, Hoshi Y, Moriguchi T, Landgraf K, Körner A, Lucas C, Brodesser S, Balogh L, Thuróczy J, Karemore G, Kuefner MS, Park EA, Rapp C, Travers JB, Röszer T. Breast milk alkylglycerols sustain beige adipocytes through adipose tissue macrophages. J Clin Invest. 2019 May 13;130:2485-2499. doi: 10.1172/JCI125646

This is incredibly cool. Thank you!
LikeLike
Amazing discovery! I look forward to more research on this topic
LikeLike
Great work!
LikeLike
Thank you ffor sharing this
LikeLike